The conceptual models were compiled by researchers in collaboration with a wide range of stakeholders from Natural Resource Management groups, universities and government agencies and based on available scientific information[2].
Click on elements of the model or select from the tabs below
Lacustrine wetlands (lakes) are dominated by open water. They are largely non-vegetated, non-riverine or non-channel systems and are not influenced by tidal waters. They are typically greater than 8 ha in size and include lakes, farm dams, reservoirs, and many other modified and highly modified systems. They often have a surrounding riparian or transitional fringe of trees. They are usually relatively deep (more than 3 m in Great Barrier Reef catchments), which limits emergent plants except around the edges, and are an important part of the landscape, providing habitat and breeding areas for a wide variety of species and many other ecosystem services. Lakes in the east coast of Queensland account for less than 2% of overall wetland area.
Lakes in Queensland, particularly in arid and semi-arid areas, are highly variable. Some are known to dry out and to support species adapted to these large changes while others stay wet for long periods and provide a refuge for many species during dry times.
The information on these pages has a focus on Queensland coastal wetlands however the principles can be used more broadly. The same principles also apply to treatment systems involved in removal of nitrogen using natural processes.
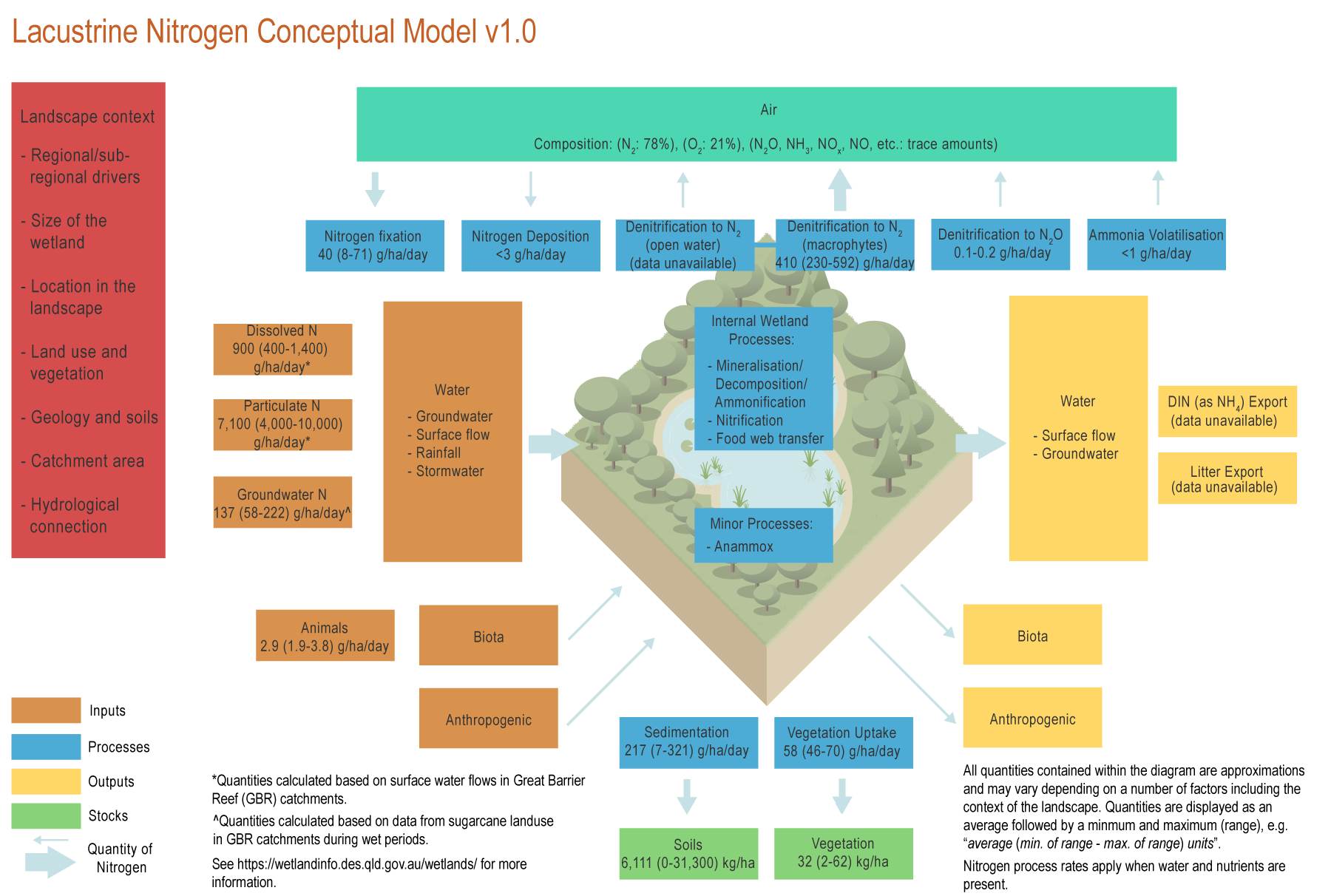
Key messages for lacustrine wetlands and nitrogen (N) processing
Nitrogen is required by wetlands for the growth of plants and animals and also as a source of energy. Additional N can be processed by lakes, providing a vital ecosystem service, however excess N can cause negative impacts on the functioning of the wetlands.[15]
As lakes are dominated by open water, which is less suitable for denitrification (the main N transformation process in wetlands)[3], they are not as effective at removing N from water as other wetland types[3], especially palustrine[3] wetlands.
N transformations in lakes are dominated by food web processes, such as incorporation into microalgae, which is consumed by zooplankton, then fish and through to other vertebrates[12][14].
Most lakes can function as a sedimentation basin, decreasing N loads associated with fine and coarse sediment, however their capacity to do so can be limited. A risk to lakes is the accumulation of excess sediment and particulate N, as this can fill the systems up.
Some lakes can have a littoral zone (the edge of the lake) with shallow water and dense aquatic plant cover (emergent, submerged and floating). These zones have high rates of denitrification, and N transformation from water to air[1][3]. Additionally, trees growing at the edges of lakes are a long-term storage for N[3].
Organic N (e.g. dead plants, dead fish) can be deposited at the bottom of the lake where it can be stored for decades or centuries[8]. Some of this deposited N can also be mineralised and released back into the water – this is not considered as a generation of N by the wetland, because the sediment was deposited from external sources.
Sometimes very high growths of weed mats (floating weeds) can form on highly nutrient rich waters of lakes. This can lead to significant drops in dissolved oxygen (DO) levels which causes the production of methane and increased ammonification, resulting in high levels of ammonium. If the weed mats are not extensive they can act as good areas for denitrification (the principle on which floating wetlands are based)[3].
Lakes have relatively high rates of N fixation compared to other wetlands, especially during nitrogen fixating algal blooms[10].
The depth of a lake is critical for N processing. Lakes that are more than about 3 m deep are not suitable for N processing because of the limited contact area between the water and the sediment and tendency to stratify in some conditions[14].
Permanent lakes process N continually. The rate at which a system processes N rich water (e.g. received from flooding) depends on the systems soil and vegetation characteristics. Ephemeral systems are usually less efficient at processing N. Permanent lakes can be fed by groundwater, including during the dry season, and by rainfall events and sporadic large floods in the wet season[13].
Lakes can have significant populations of birds and other animals. These inputs of N (e.g. droppings and dead animals) to the wetland are relatively small, but can be important at a local scale, for instance, in shallow ponds[9].
Cyanobacterial (blue-green algae) blooms in lakes are a natural process that can be exacerbated by increases in N loads[4]. Increases in N loads can also result in increases in macrophyte growth. Increases in organic matter production by algae or macrophytes can cause a decrease in DO in the bottom of the lake and the liberation of ammonium leading to fish kills[5].
The condition or state of lakes can affect how the system processes N. High levels of weeds, significant amounts of sediment, anaerobic conditions (no oxygen) and changes to hydrology can have large impacts on the efficiencies of the system[5].
Lakes within the Great Barrier Reef (GBR) catchments are either subtropical or tropical and are relatively shallow (less than 20 m)[14]. They are easily mixed and result in a homogeneous water column throughout the year with permanent or sporadic high turbidity[14].
Lakes often produce N gas (N2) from denitrification[16]. If a lake contains high N loads, it can generate a higher proportion of N2O[7], a greenhouse gas, and NH3, which is toxic to aquatic fauna[11], depending on temperature and pH.
The processing of N by lakes is affected by the concentration of N entering the wetland, with denitrification at higher N concentrations[1].
Lakes are generally located in low-lying areas and can be natural or artificially created. Natural lacustrine wetlands are usually floodplain lakes with intermittent connection to the main river channel during flood events. Due to their location in the catchment, they receive large quantities of nutrients, mostly via surface runoff in the wet season and groundwater in the dry season[6]. See Regional, subregional and landscape context page for further information.
References
^ab Adame, MF, Roberts, ME, Hamilton, DP, Ndehedehe, CE, Lu, J, Griffiths, M, Curwen, G & Ronan, M (2019), 'Tropical coastal wetlands ameliorate nitrogen exports during floods', Frontiers in Marine Science, vol. 6, 1-14.
^ Adame, MF, Vilas, MP, Franklin, H, Garzon-Garcia, A, Hamilton, D, Ronan, M & Griffiths, M (2021), 'A conceptual model of nitrogen dynamics for the Great Barrier Reef catchments', Marine Pollution Bulletin. [online], vol. 173PA. Available at: https://www.sciencedirect.com/science/article/pii/S0025326X21009437.
^abcdef Adame, MF, Waltham, NJ, Iram, N, Farahani, BS, Salinas, C, Burford, M & Ronan, M (8 July 2021), 'Denitrification within the sediments and epiphyton of tropical macrophyte stands', Inland Waters. [online], pp. 1-10. Available at: https://www.tandfonline.com/doi/full/10.1080/20442041.2021.1902214 [Accessed 31 July 2021].
^ Burford, MA, Mcneale, KL & Mckenzie-Smith, FJ (November 2006), 'The role of nitrogen in promoting the toxic cyanophyte Cylindrospermopsis raciborskii in a subtropical water reservoir', Freshwater Biology. [online], vol. 51, no. 11, pp. 2143-2153. Available at: http://doi.wiley.com/10.1111/j.1365-2427.2006.01630.x [Accessed 30 October 2020].
^ab Cary, PR & Weerts, PGJ (June 1983), 'Growth of Salvinia molesta as affected by water temperature and nutrition I. Effects of nitrogen level and nitrogen compounds', Aquatic Botany. [online], vol. 16, no. 2, pp. 163-172. Available at: https://linkinghub.elsevier.com/retrieve/pii/0304377083900918 [Accessed 2 November 2020].
^ Connolly, NM, Pearson, RG, Loong, D, Maughan, M & Brodie, J (2015), 'Water quality variation along streams with similar agricultural development but contrasting riparian vegetation', Agriculture, Ecosystems and Environment. [online], vol. 213, no. x, pp. 11-20. Available at: http://dx.doi.org/10.1016/j.agee.2015.07.007.
^ Deemer, BR, Harrison, JA, Li, S, Beaulieu, JJ, DelSontro, T, Barros, N, Bezerra-Neto, JF, Powers, SM, dos Santos, MA & Vonk, JA (1 November 2016), 'Greenhouse Gas Emissions from Reservoir Water Surfaces: A New Global Synthesis', BioScience. [online], vol. 66, no. 11, pp. 949-964. Available at: https://academic.oup.com/bioscience/article/66/11/949/2754271 [Accessed 30 October 2020].
^ Haberle, SG (2018), 'A 23,000-yr pollen record from Lake Euramoo, Wet Tropics of NE Queensland, Australia', Quaternary Research, vol. 64, no. 2005, pp. 343-356.
^ Hahn, S, Bauer, S & Klaassen, M (2008), 'Quantification of allochthonous nutrient input into freshwater bodies by herbivorous waterbirds', Freshwater Biology, vol. 53, pp. 181-193.
^ Howarth, RW, Marino, R, Lane, J & Cole, JJ (1988), 'Nitrogen fixation in freshwater, estuarine, and marine ecosystems. 1. Rates and importance', Limnology and Oceanography, vol. 33, pp. 669-687.
^ Ip, YK, Chew, SF, Wilson, JM & Randall, DJ (17 August 2004), 'Defences against ammonia toxicity in tropical air-breathing fishes exposed to high concentrations of environmental ammonia: a review', Journal of Comparative Physiology B. [online] Available at: http://link.springer.com/10.1007/s00360-004-0445-1 [Accessed 30 July 2021].
^ Jardine, TD, Pusey, BJ, Hamilton, SK, Pettit, NE, Davies, PM, Douglas, MM, Sinnamon, V, Halliday, IA & Bunn, SE (2012), 'Fish mediate high food web connectivity in the lower reaches of a tropical floodplain river', Oecologia, vol. 168, pp. 829-838.
^ Karim, F, Kinsey-henderson, A, Wallace, J, Arthington, AH & Pearson, RG (2012), Modelling wetland connectivity during overbank flooding in a tropical floodplain in north Queensland, Australia, vol. 2723, pp. 2710-2723.
^abcd Lewis, WM (1996), 'Tropical lakes : how latitude makes a difference', Perspectives in Tropical Limnology, p. 43, SPB Academic Publishing, Amsterdam, The Netherlands, eds. F Schiemer & K T Boland.
^ Mitsch, WJ & Gosselink, J (2015), Wetlands, p. 213, Wiley, New Jersey, USA.
^ Sturm, K, Yuan, Z, Gibbes, B, Werner, U & Grinham, A (30 September 2014), 'Methane and nitrous oxide sources and emissions in a subtropical freshwater reservoir, South East Queensland, Australia', Biogeosciences. [online], vol. 11, no. 18, pp. 5245-5258. Available at: https://bg.copernicus.org/articles/11/5245/2014/ [Accessed 30 October 2020].
Last updated: 9 August 2021
This page should be cited as:
Department of Environment, Science and Innovation, Queensland (2021) Lacustrine, WetlandInfo website, accessed 30 August 2024. Available at: https://wetlandinfo.des.qld.gov.au/wetlands/ecology/processes-systems/nitrogen-concept-model/lacustrine/





 — Department of Environment, Science and Innovation
— Department of Environment, Science and Innovation